
1º BAT
4º ESO Interpretación de cortes geológicos
Os dejo un pequeño powerpoint realizado por los profesores de la Universidad de Alicante que puede ayudaros a comprender los corte geológicos.
Os dejo un pequeño powerpoint realizado por los profesores de la Universidad de Alicante que puede ayudaros a comprender los corte geológicos.

Os adjunto aquí el boletín 2 de problemas de genética. La ciencia es más que un simple conjunto de conocimientos: es una
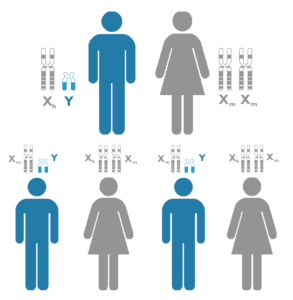
Lo primero que vamos a ver es como se determina el sexo de los descendientes en diferentes especies animales. Existen cuatro posibilidades en la naturaleza:
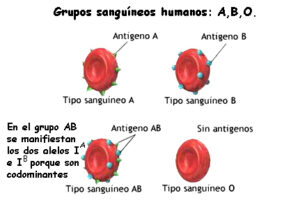
Además de las leyes anteriores en el que los híbridos muestran el mismo genotipo que uno de los parentales, lo que se llama dominancia total

Esquemas utilizados en la clase do hoy ¡Hemos encontrado el secreto de la Vida! En la mañana del 28 de Febrero de 1953, Francis
Nos obligan a avisaros de que usamos cookies. Si pulsas ok, estaremos muy contentos Ok